kromosomer
Definition - hvad er kromosomer?
En celles genetiske sammensætning opbevares i form af DNA (deoxyribonukleinsyre) og dens baser (adenin, thymin, guanin og cytosin). I alle eukaryote celler (dyr, planter, svampe) er dette til stede i cellekernen i form af kromosomer. Et kromosom består af et enkelt, sammenhængende DNA-molekyle, som er knyttet til visse proteiner.
Navnet kromosom er afledt af græsk og kan groft oversættes til "farvelegeme". Dette navn stammer fra det faktum, at meget tidligt i cytologiens historie (1888) lykkedes det forskerne med at farve dem ved hjælp af specielle basale farvestoffer og identificere dem i et lysmikroskop. De er imidlertid kun virkelig synlige på et bestemt tidspunkt i cellecyklussen, mitose (meiose i kimceller), når kromosomet er særlig tæt (kondenseret).

Hvordan er kromosomer struktureret?
Hvis hele DNA-dobbelthelixen af en celle, dvs. omkring 3,4 x 109 basepar, skulle kobles sammen, ville dette resultere i en længde på over en meter. Den samlede længde af alle tilsatte kromosomer er kun ca. 115 um. Denne forskel i længde forklares med den meget kompakte struktur af kromosomerne, hvor DNA'et er viklet eller spiralformet flere gange på en meget specifik måde.
Histoner, en speciel form for proteiner, spiller en vigtig rolle i dette. Der er i alt 5 forskellige histoner: H1, H2A, H2B, H3 og H4. To af de sidste fire histoner kombineres for at danne en cylindrisk struktur, oktameren, omkring hvilken den dobbelte helix vikles omkring to gange (= superhelix). H1 binder sig til denne struktur for at stabilisere den.
Dette kompleks af DNA, oktamer og H1 kaldes et nukleosom. Flere af disse nukleosomer er nu "som en streng med perler" med relativt korte intervaller (10-60 basepar) den ene bag den anden. Sektionerne mellem kromosomerne er kendt som spacer-DNA. De enkelte nukleosomer kommer nu i kontakt igen via H1, hvilket skaber en yderligere spiralform og dermed også en kompression.
Den resulterende streng er igen til stede i løkker, som er stabiliseret af en rygrad lavet af sure ikke-histonproteiner, også kendt som Hertone. Disse løkker er til gengæld spiraler, der er stabiliseret af proteiner, hvilket resulterer i det sidste trin i kompression. Imidlertid forekommer denne høje grad af kompression kun i forbindelse med celledeling under mitose.
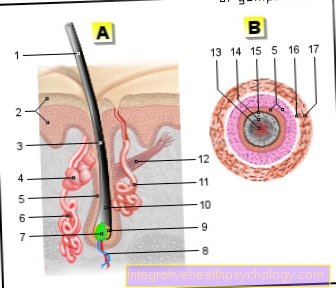
I denne fase kan du også se den karakteristiske form af kromosomerne, der er sammensat af to kromatider. Det sted, hvor disse er forbundet, kaldes centromeren. Det deler hvert metafase-kromosom i to korte og to lange arme, også kaldet p og q-arme.
Hvis centromeren ligger nogenlunde i midten af kromosomet, kaldes det et metacentrisk kromosom, hvis det helt ligger i en af enderne af et akrocentrisk kromosom. De imellem kaldes submetacentriske kromosomer. Disse forskelle, som allerede kan ses under lysmikroskopet sammen med længden, tillader en indledende klassificering af kromosomerne.
Hvad er telomererne?
Telomerer er enderne af kromosomerne med gentagne sekvenser (TTAGGG). Disse bærer ikke nogen relevant information, men tjener snarere til at forhindre tab af mere relevante DNA-sektioner. Ved hver celledeling går en del af kromosomet tabt gennem mekanismen for DNA-replikation.
Så telomererne er på en måde en buffer, der forsinker det punkt, hvor cellen mister vigtig information gennem opdeling. Hvis telomererne i en celle er mindre end 4.000 basepar i længde, initieres programmeret celledød (apoptose). Dette forhindrer spredning af defekt genetisk materiale i organismen. Et par celler har telomeraser, enzymer, der er i stand til at forlænge telomererne igen.
Ud over de stamceller, hvorfra alle andre celler stammer, er dette kimceller og visse celler i immunsystemet. Derudover findes telomeraser også i kræftceller, hvorfor man taler om udødeliggørelse i denne celle.
Læs alt om emnet her: Telomerer - Anatomi, funktion og sygdomme
Hvad er kromatin?
Chromatin henviser til hele indholdet af en cellekerne, der kan farves med en base. Derfor inkluderer udtrykket ud over DNA også visse proteiner, f.eks. Histoner og hertoner (se struktur) samt visse RNA-fragmenter (hn og snRNA).
Afhængigt af fasen i cellecyklussen eller afhængig af genetisk aktivitet, er dette materiale tilgængeligt i forskellige densiteter. Den tættere form kaldes heterochromatin. For at gøre det lettere at forstå, kunne man derfor betragte det som en "opbevaringsform" og her igen skelne mellem konstitutivt og fakultativt heterochromatin.
Konstitutivt heterochromatin er den mest tætte form, der er til stede i dets højeste niveau af kondensation i alle faser af cellecyklussen. Det udgør omkring 6,5% af det humane genom og er hovedsageligt beliggende nær centromererne og enderne af kromosomarme (telomerer) i lille grad, men også andre steder (hovedsageligt kromosom 1, 9, 16, 19 og Y). Derudover er det meste af det konstitutive heterochromatin lokaliseret nær kernemembranen, dvs. på kanterne af cellekernen. Mellemrummet er forbeholdt den aktive kromatin, euchromatinet.
Facultativt heterochromatin er lidt mindre tæt og kan aktiveres og deaktiveres efter behov eller afhængigt af udviklingsstadiet. Et godt eksempel på dette er det andet X-kromosom i kvindelige karyotyper. Da et X-kromosom dybest set er nok til, at cellen kan overleve, hvilket i sidste ende er tilstrækkeligt for mænd, deaktiveres en af de to i den embryonale fase. det deaktiverede X-kromosom er kendt som Barr-kroppen.
Kun under celledeling kondenseres den i forbindelse med mitose fuldstændigt, hvorved den når sin højeste kompression i metafasen. Da de forskellige gener imidlertid læses forskelligt ofte - når alt kommer til alt kræves ikke hvert protein i samme mængde på alle tidspunkter - sondres her mellem aktiv og inaktiv euchromatin.
Læs mere om dette under: Chromatin
Haploide kromosomer
Haploid (Gr. Haploos = enkelt) betyder, at alle kromosomer i en celle er til stede individuelt, dvs. ikke i par (diploid), som normalt er tilfældet. Dette er den naturlige tilstand af alle æg- og sædceller, hvor de to identiske kromatider ikke oprindeligt er adskilt som en del af den første meiose, men snarere alle kromosompar adskilles først.
Som et resultat, efter den første meiose, har dattercellerne hos mennesker kun 23 i stedet for de sædvanlige 46 kromosomer, hvilket svarer til halvdelen af det haploide sæt kromosomer. Da disse datterceller stadig har en identisk kopi af hvert kromosom bestående af 2 kromosomer, kræves den anden meiose, hvor de to kromatider er adskilt fra hinanden.
Polytenkromosomer
Et polytenkromosom er et kromosom, der består af et stort antal genetisk identiske kromatider. Da sådanne kromosomer er lette at se under en lavere forstørrelse, omtales de undertiden som gigantiske kromosomer. Forudsætningen herfor er endoreplikation, hvori kromosomerne multipliceres flere gange i cellekernen, uden at celledeling finder sted.
Hvad er kromosomernes funktioner?
Kromosomet, som den organisatoriske enhed for vores genom, bruges primært til at sikre, at det duplikerede genom fordeles jævnt mellem dattercellerne under celledeling. For at gøre dette er det værd at se nærmere på mekanismerne for celledeling eller cellecyklus:
Cellen tilbringer det meste af cellecyklussen i interfasen, hvilket betyder hele den periode, hvor cellen ikke straks er ved at dele sig. Dette er igen opdelt i G1-, S- og G2-faser.
Gl-fasen (G for gap, dvs. gap) følger straks celledeling. Her øges cellen i størrelse igen og udfører generelle metaboliske funktioner.
Herfra kan det også skifte til G0-fasen. Dette betyder, at det skifter til et trin, der ikke længere er i stand til at dele sig, og i normale tilfælde også ændrer sig meget for at udføre en meget specifik funktion (celledifferentiering). For at udføre disse opgaver læses meget specifikke gener mere intenst, andre mindre eller slet ikke.
Hvis det ikke er nødvendigt med en sektion af DNA i lang tid, er den ofte placeret i de dele af kromosomerne, der har været tætpakket i lang tid (se kromatin). På den ene side har dette formålet at spare plads, men ud over de andre mekanismer til genregulering er det også en ekstra beskyttelse mod at blive læst ved et uheld. Imidlertid er det også observeret, at under meget specifikke betingelser kan differentierede celler fra GO-fasen genindtræde i cyklussen.
G1-fasen efterfølges af S-fasen, dvs. den fase, hvor nyt DNA syntetiseres (DNA-replikation). Her skal hele DNA være i sin løseste form, dvs. alle kromosomer er fuldstændigt ikke viklet (se struktur).
Ved afslutningen af syntesefasen duplikeres hele det genetiske materiale i cellen. Da kopien stadig er knyttet til det originale kromosom via centromeren (se struktur), taler man ikke om en duplikation af kromosomerne.
Hvert kromosom består nu af to kromatider i stedet for et, så det senere kan antage den karakteristiske X-form under mitose (strengt taget gælder X-formen kun for metacentriske kromosomer). I den efterfølgende G2-fase finder den øjeblikkelige forberedelse til celledeling sted. Dette inkluderer også en detaljeret kontrol af replikationsfejl og strengbrud, som kan repareres om nødvendigt.
Der er dybest set to typer celledeling: mitose og meiose. Med undtagelse af kimcellerne opstår alle celler i en organisme gennem mitose, hvis eneste opgave er dannelsen af to genetisk identiske datterceller.
Meiosis har derimod formålet at generere genetisk forskellige celler:
I et første trin opdeles de tilsvarende (homologe), men ikke identiske kromosomer. Kun i det næste trin er kromosomerne, der består af to identiske chromatider, adskilt og igen fordelt til to datterceller, så til sidst opstår fire kimceller med forskelligt genetisk materiale fra en precursorcelle.
Kromosomernes form og struktur er essentiel for begge mekanismer: Specielle "proteintråde", det såkaldte spindelapparat, fastgøres til de stærkt kondenserede kromosomer og trækker kromosomerne i en finreguleret proces fra midtplanet (ækvatorplan) til de modsatte poler i cellen omkring en for at sikre jævn distribution. Selv små ændringer i kromosomernes mikrostruktur kan have alvorlige konsekvenser.
I alle pattedyr bestemmer forholdet mellem kønskromosomer X og Y også afkommets køn. Grundlæggende afhænger det hele af, om sædcellen, der forenes med æggecellen, bærer et X- eller Y-kromosom. Da begge former for sædceller altid produceres i nøjagtigt samme omfang, er sandsynligheden altid afbalanceret for begge køn. Dette tilfældige system garanterer en mere jævn kønsfordeling end f.eks. Med miljøfaktorer som temperatur.
Find ud af mere om emnet: Cellekernedeling
Hvordan overføres generne via kromosomerne?
I dag ved vi, at træk arves via gener, der er lagret i cellerne i form af DNA. Disse er igen delt i 46 kromosomer, på hvilke de 25.000-30000 humane gener er fordelt.
Ud over selve ejendommen, der kaldes fænotype, findes der også den genetiske modstykke, der kaldes genotypen. Det sted, hvor et gen er på et kromosom, kaldes et locus. Da mennesker har det dobbelte af hvert kromosom, forekommer hvert gen også to gange. Den eneste undtagelse herfra er X-kromosomgenerne hos mænd, da Y-kromosomet kun bærer en brøkdel af den genetiske information, der findes på X-kromosomet.
Forskellige gener, der er på det samme lokus, kaldes alleler. Der er ofte mere end to forskellige alleler på et lokus. Man taler derefter om polymorfisme. En sådan allel kan simpelthen være en ufarlig variant (normal variant), men også patologiske mutationer, som kan være udløseren til en arvelig sygdom.
Hvis mutationen af et enkelt gen er tilstrækkeligt til at ændre fænotypen, taler man om monogen eller Mendelian arv. Mange af de arvelige træk arves imidlertid gennem flere interagerende gener og er derfor meget vanskeligere at studere.
Da mor og far hver videregiver en af deres to gener til barnet i en Mendelian arv, er der altid fire mulige kombinationer i den næste generation, hvorved disse også kan være de samme i forhold til en egenskab. Hvis begge alleler hos et individ har den samme effekt på fænotypen, er individet homozygot i forhold til denne egenskab, og karakteristikken udtrykkes tilsvarende fuldt ud.
Heterozygoter har to forskellige alleler, der kan interagere med hinanden på forskellige måder: Hvis en allel er dominerende over en anden, undertrykker den fuldstændigt dens udtryk, og den dominerende egenskab bliver synlig i fænotypen. Den undertrykte allel kaldes recessiv.
I tilfælde af en kodominant arv kan begge alleler udtrykke sig upåvirket af hinanden, mens der i tilfælde af en mellemarv er en blanding af begge egenskaber. Et godt eksempel på dette er AB0-blodgruppesystemet, hvor A og B er co-dominerende med hinanden, men 0 er dominerende over hinanden.
Hvad er det normale sæt kromosomer hos mennesker?
Humane celler har 22 kønsuafhængige par kromosomer (autosomer) og to kønskromosomer (gonosomer), så i alt 46 kromosomer udgør et sæt kromosomer.
Autosomer kommer normalt parvis. Kromosomerne i et par er ens i form og sekvens af gener og omtales derfor som homologe. De to X-kromosomer hos kvinder er også homologe, mens mænd har et X- og Y-kromosom. Disse adskiller sig i form og antal gener, der er til stede, så man ikke længere kan tale om homologi.
Kimceller, dvs. æg og sædceller, har kun halvdelen af kromosomsættet på grund af meiose, nemlig 22 individuelle autosomer og et gonosom. Da kimcellerne smelter sammen under befrugtning og sommetider bytter hele segmenter (crossover), oprettes en ny kombination af kromosomer (rekombination). Alle kromosomer kaldes sammen karyotypen, der med få undtagelser (se kromosomafvigelser) er identisk i alle individer af samme køn.
Her kan du finde ud af alt om emnet: Mitose - simpelthen forklaret!
Hvorfor er der altid par kromosomer?
Grundlæggende kan dette spørgsmål besvares med en sætning: Fordi det har vist sig at være fordelagtigt. Tilstedeværelsen af kromosompar og princippet om rekombination er essentielle for arv i form af seksuel reproduktion. På denne måde kan et helt nyt individ komme ud af det enkelte menneskes genetiske materiale ved en tilfældighed.
Dette system forøger enormt forskellige egenskaber inden for en art og sikrer, at det kan tilpasse sig ændrede miljøforhold meget hurtigere og mere fleksibelt, end det kun ville være muligt gennem mutation og selektion.
Det dobbelte sæt kromosomer har også en beskyttende virkning: hvis en mutation af et gen ville føre til en funktionssvigt, er der stadig en slags "sikkerhedskopi" i det andet kromosom. Dette er ikke altid nok til, at organismen kompenserer for fejlen, især hvis den muterede allel er dominerende, men det øger chancen for det. Derudover overføres mutationen ikke automatisk til alle afkom, hvilket igen beskytter arten mod for radikale mutationer.
Hvad er en kromosommutation?
Genetiske defekter kan opstå fra ioniserende stråling (f.eks. Røntgenstråler), kemiske stoffer (f.eks. Benzopyren i cigaretrøg), visse vira (f.eks. HP-vira) eller, med en lav sandsynlighed, de kan også opstå rent tilfældigt. Der er ofte flere faktorer involveret i dens udvikling. I princippet kan sådanne ændringer forekomme i alt kropsvæv, men af praktiske grunde er analysen sædvanligvis begrænset til lymfocytter (en speciel type immuncelle), fibroblaster (bindevævsceller) og knoglemarvsceller.
En kromosommutation er en stor strukturel ændring i individuelle kromosomer.Fraværet eller tilsætningen af hele kromosomer ville på den anden side være et genom- eller ploidiemutation, medens udtrykket genmutation refererer til relativt små ændringer inden for et gen. Udtrykket kromosomafvigelse (latin aberrare = at afvige) er noget bredere og inkluderer alle ændringer, der kan detekteres med et lysmikroskop.
Mutationer kan have meget forskellige effekter:
- Stille mutationer, dvs. mutationer, hvor ændringen ikke har nogen indflydelse på individet eller deres afkom, er temmelig atypiske for kromosomafvigelser og findes oftere i området med gen- eller punktmutationer.
- En tab af funktionsmutation er, når mutationen resulterer i et fejlagtigt og derfor funktionelt protein eller slet intet protein.
- Såkaldte gain-of-function mutationer ændrer typen af effekt eller mængden af proteiner produceret på en sådan måde, at der opstår helt nye effekter. På den ene side er dette en afgørende mekanisme for evolution og dermed for overlevelse af en art eller fremkomsten af nye arter, men på den anden side, som i tilfældet med Philadelphia-kromosomet, kan det også yde et afgørende bidrag til udviklingen af kræftceller.
Den bedst kendte af de forskellige former for kromosomafvigelser er sandsynligvis de numeriske afvigelser, hvor individuelle kromosomer kun er til stede en gang (monosomi) eller endda tredobbelt (trisomi).
Hvis dette kun gælder et enkelt kromosom, kaldes det aneuploidi, og hele kromosomsættet påvirkes af polyploidi (tri- og tetraploidi). I de fleste tilfælde opstår denne maldistribution i løbet af kimcelleudvikling på grund af ikke-adskillelse (nondisjunction) af kromosomerne under celledeling (meiose). Dette fører til en ujævn fordeling af kromosomerne mellem dattercellerne og dermed til numerisk afvigelse hos det udviklende barn.
Monosomier af ikke-kønskromosomer (= autosomer) er uforenelige med livet og forekommer derfor ikke hos levende børn. Med undtagelse af trisomier 13, 18 og 21 fører autosomale trisomier næsten altid til spontane aborter.
Under alle omstændigheder er der i modsætning til afvigelser fra kønskromosomerne, som også kan være iøjnefaldende, altid alvorlige kliniske symptomer, og som regel mere eller mindre udtalt ydre abnormiteter (dysmorfismer).
En sådan maldistribution kan også forekomme senere i livet med mitotisk celledeling (alle celler undtagen kimceller). Da der også er uændrede celler ud over de berørte celler, taler man om en somatisk mosaik. Med somatisk (græsk soma = krop) menes alle celler, der ikke er kimceller. Da kun en lille del af kroppens celler påvirkes, er symptomerne som regel meget mildere. Derfor mosaiktyper ofte opdages i lang tid.
Her kan du finde ud af alt om emnet: Kromosommutation
Hvad er kromosomafvigelse?
Strukturel kromosomafvigelse svarer stort set til definitionen af kromosommutation (se ovenfor). Hvis mængden af genetisk materiale forbliver den samme og simpelthen distribueres forskelligt, taler man om en afbalanceret afvigelse.
Dette sker ofte via translokation, dvs. overførsel af et kromosomsegment til et andet kromosom. Hvis det er en udveksling mellem to kromosomer, taler man om gensidig translokation. Da kun ca. 2% af genomet er påkrævet for at producere proteiner, er sandsynligheden meget lav for, at et sådant gen er ved brudpunktet og derved mister sin funktion eller er nedsat i det. Derfor går en sådan afbalanceret afvigelse ofte upåagtet hen og overføres over flere generationer.
Dette kan imidlertid føre til en forkert fordeling af kromosomer under udviklingen af kimcellerne, hvilket kan resultere i infertilitet, spontane aborter eller afkom med en ubalanceret afvigelse.
Imidlertid kan en ubalanceret afvigelse også forekomme spontant, dvs. uden en familiehistorie. Sandsynligheden for, at et barn bliver født i live med ubalanceret afvigelse, afhænger meget af de berørte kromosomer og varierer mellem 0 og 60%. Dette fører til tab (= sletning) eller duplikation (= duplikering) af et kromosomsegment. I denne sammenhæng taler man om delvise mono- og trisomier.
I nogle tilfælde forekommer disse sammen i to forskellige regioner, hvor delvis monosomi normalt er mere afgørende for udseendet af kliniske symptomer. Dette er fremtrædende eksempler på en sletning Cat Scream Syndrome og Wolf-Hirschhorn Syndrome.
Man taler om en mikrodeletion, når ændringen ikke længere kan bestemmes med lysmikroskopet, dvs. når det er et spørgsmål om tabet af en eller et par gener. Dette fænomen anses for at være årsagen til Prader-Willi syndrom og Angelman syndrom og er tæt forbundet med udviklingen af retionoblastom.
Robertson-translokationen er et specielt tilfælde:
To acrocentriske kromosomer (13, 14, 15, 21, 22) forenes ved deres centromere og danner, efter at de har mistet de korte arme, et enkelt kromosom (se struktur). Selvom dette resulterer i et reduceret antal kromosomer, kaldes dette en afbalanceret afvigelse, da tabet af de korte arme i disse kromosomer let kan kompenseres for. Også her ses virkningerne ofte kun i efterfølgende generationer, da der er en meget stor sandsynlighed for aborter eller levende børn med en trisomi.
Hvis der er to pauser i et kromosom, kan det ske, at det mellemliggende segment roteres 180 ° og inkorporeres i kromosomet. Denne proces, kendt som inversion, er kun ubalanceret, hvis knækpunktet ligger inden for et aktivt gen (2% af det samlede genetiske materiale). Afhængigt af om centromeren er inden i eller uden for det inverterede segment, er det en peri- eller paracentrisk inversion. Disse ændringer kan også bidrage til ujævn fordeling af det genetiske materiale på kimcellerne.
Ved paracentrisk inversion, hvor centromeren ikke er i det inverterede segment, kan kimceller med to eller ingen centromer også vises. Som et resultat går det tilsvarende kromosom tabt under de allerførste celledelinger, hvilket næsten helt sikkert fører til en spontanabort.
Indsættelse henviser til inkorporering af et kromosomfragment andetsteds. Også her påvirkes afkom primært på en lignende måde. Et ringkromosom kan forekomme især efter sletning af endestykker. Typen og størrelsen af sekvenserne er afgørende for sværhedsgraden af symptomerne. Derudover kan dette føre til forkerte fordelinger og dermed igen til mosaiktyper i kroppens celler.
Hvis metaphase-kromosomet adskiller sig forkert under celledeling, kan der være isochromosomer. Dette er to nøjagtigt de samme kromosomer, der kun består af lange eller kun korte arme. I tilfælde af X-kromosom kan dette manifestere sig som et Ulrich-Turner-syndrom (monosomi X).
Læs mere om dette emne: Kromosomafvigelse
Trisomi 21
Trisomi 21, bedre kendt som Downs syndrom, er uden tvivl den mest almindelige numeriske kromosomale afvigelse blandt levende fødsler, hvor mænd påvirkes lidt oftere (1.3: 1).
Sandsynligheden for, at trisomi 21 kan forekomme, afhænger af forskellige demografiske faktorer, såsom gennemsnitsalderen ved fødslen af mødrene, og varierer lidt fra region til region.
95% af trisomien 21 opstår som et resultat af en opdelingsfejl i sammenhæng med meiose (kimcelledeling), nemlig nondisjunktion, dvs. manglen på at adskille søsterchromatiderne.
Disse benævnes fri trisomier og opstår 90% i moderen, 5% i faderen og yderligere 5% i det embryonale genom.
Yderligere 3% er resultatet af ubalancerede translokationer enten på kromosom 14 eller som 21; 21 translokation, hvilket skaber et normalt og et dobbelt kromosom 21. De resterende 2% er mosaiktyper, hvor trisomien ikke opstod i kimceller og derfor ikke påvirker alle kropsceller. Mosaiktyper er ofte så milde, at de kan forblive helt uopdagede i lang tid.
Under alle omstændigheder skal en kromosomundersøgelse udføres for at skelne den symptomatisk identiske frie trisomi fra den muligvis nedarvede translokationstrisomi. En familiehistorie fra de foregående generationer kan derefter følge.
Er du interesseret i dette emne? Læs den næste artikel om dette: Trisomi 21
Trisomi 13
Trisomi 13 eller Patau syndrom har en frekvens på 1: 5000 og er meget sjældnere end Downs syndrom. Årsagerne (gratis trisomier, translokationer og mosaiktyper) og deres procentvise fordeling er dog stort set identiske.
I teorien kunne næsten alle tilfælde diagnosticeres prenatalt ved hjælp af ultralyd eller PAPP-A-testen. Da PAPP-A-testen ikke nødvendigvis er en del af de rutinemæssige undersøgelser, diagnosticeres omkring 80% af tilfældene i Centraleuropa før fødslen.
En vækstrest, en bilateral spalteleppe og gane og usædvanligt små øjne (mikrophthalmia) kan allerede ses ved ultralyd. Derudover er der normalt misdannelser i forhjernen og ansigtet med forskellige sværhedsgrader (holoprosencephaly).
Mens cerebrale halvkugler i lobær form er næsten fuldstændigt adskilt, og der dannes laterale ventrikler, i semi-lobar form, adskilles ofte kun den bageste del af hjernen, og de laterale ventrikler mangler. I den mest alvorlige form, den alobare form, er der ingen adskillelse af de cerebrale halvkugler.
Børn med halv- eller alobar form dør normalt umiddelbart efter fødslen. Efter en måned er dødeligheden ca. 50% af levende fødsler. Indtil 5-årsalderen stiger dødeligheden fra trisomi 13 til 90%. På grund af misdannelser i hjernen forbliver de syge i de fleste tilfælde sengeliggende i livet og kan ikke tale, hvorfor de er afhængige af fuld pleje. Derudover kan der også være omfattende fysiske manifestationer af Trismoie 13.
Læs mere om emnet på: Trisomi 13 i det ufødte barn
Trisomi 16
Grundlæggende er trisomi 16 den mest almindelige trisomi (ca. 32% af alle trisomier), men levende børn med trisomi 16 er meget sjældne. Generelt forekommer levende fødsler kun i partielle trisomier eller mosaiktyper. Blandt trisomierne er det oftest ansvarlig for dødfødsler: 32 ud af 100 aborter på grund af kromosomafvigelser kan spores tilbage til denne form for trisomi.
Derfor er der hovedsageligt dokumenteret prenatal, dvs. prenatal, identificerbare egenskaber. Bemærkelsesværdigt her er forskellige hjertedefekter, nedsat vækst, en enkelt navlens arterie (ellers dobbelt) og øget nakkets gennemsigtighed, hvilket forklares med væskeansamling på grund af det endnu ikke fuldt udviklede lymfesystem og den øgede elasticitet af huden i dette område. Derudover regresserer ofte ikke den fysiologiske navlebrok, dvs. den midlertidige forskydning af en stor del af tarmen gennem navlen til ydersiden, hvilket er kendt som en omphalocele- eller navlestrengsbrud.
En flexionskontraktur med krydsede fingre kan også ofte påvises ved ultralyd. I de få levende fødsler bemærkes generaliseret muskelhypotension, dvs. generel muskelsvaghed. Dette fører til drikkesvaghed og kan sikre, at barnet skal fodres kunstigt. Firefinger-furen, der er så karakteristisk for trisomier, forekommer ofte. Også her er hyppigheden af forekomst af trisomien direkte relateret til moderens alder.
Trisomi 18
Edwards syndrom, dvs. trisomi 18, forekommer med en frekvens på 1: 3000. Ved prænatal diagnostik er det det samme som med Patau-syndromet: Også her ville de samme undersøgelser give mulighed for, at alle patienter findes helt før fødslen. Årsagerne og fordelingen heraf skal sammenlignes med andre trisomier (se trisomi 21).
Derudover findes der i trisomi 18 også delvis trisomier, som ligesom mosaiktyperne fører til meget mildere kliniske kurser. De tilknyttede dysmorfismer er også ekstremt karakteristiske for Edwards syndrom: Ved fødslen har patienter en stærkt reduceret kropsvægt på 2 kg (normal: 2,8-4,2 kg), et tilbagevendende bredt pande, en generelt underudviklet nedre halvdel af ansigtet med en lille mundåbning , smalle øjenlåg og roterede bagud, formændrede ører (fauns øre). Derudover er bagsiden af hovedet usædvanligt veludviklet til en nyfødt. Ribbenene er usædvanligt smalle og skrøbelige. Nyfødte har også en permanent spænding (tone) i hele muskulaturen, som dog forsvinder efter de første par uger hos de overlevende.
Et andet karakteristisk træk er krydsningen af 2. og 5. fingre over 3. og 4. med det samlede antal fingre, der er hamret ind, mens fødderne er usædvanligt lange (forløbne), har en særlig udtalt hæl, forkrøblede tånegle og en tilbagestående stortå.
Alvorlige organdeforfunktioner er almindelige og forekommer normalt i kombination: hjerte- og nyredefekter, foldfoldning (malrotation) i tarmen, adhæsioner i bughinden (mesenterium kommune), en okklusion af spiserøret (øsofagus atresi) og mange flere.
På grund af disse misdannelser er dødeligheden omkring 50% inden for de første 4 dage, kun omkring 5-10% lever for at være over et år. Overlevelse i voksen alder er den absolutte undtagelse. Under alle omstændigheder er en intellektuel handicap meget udtalt og kan ikke tale, er sengeliggende og inkontinent, så helt afhængig af hjælp udefra.
For mere detaljeret information om trisomi 18, bedes du læse vores detaljerede artikel om emnet:
- Trisomi 18 (Edwards syndrom)
- Trisomi 18 i det ufødte barn
Trisomi X
Trisomi X er den mest iøjnefaldende form for numerisk kromosomafvigelse, udseendet af de berørte, som logisk set alle er kvindelige, adskiller sig ikke meget fra andre kvinder. Nogle er mærkbare, fordi de er særlig høje og har noget "fyldige" ansigtstræk. Psykisk udvikling kan også være stort set normal og spænder fra normal grænse til mild psykisk handicap.
Imidlertid er dette intelligensunderskud lidt mere alvorligt end med de andre trisomier i sexkromosomerne (XXY og XYY). Med en frekvens på 1: 1000 er det faktisk ikke så sjældent, men da trisomien normalt ikke er forbundet med klinisk signifikante symptomer, vil størstedelen af kvinder med sygdommen sandsynligvis aldrig blive diagnosticeret i hele deres liv.
Transportører opdages for det meste ved en tilfældighed under en familiekontrol eller under prenatal diagnostik. Fertiliteten kan være svagt reduceret, og frekvensen af afvigelser i kønkromosomer i den næste generation kan øges lidt, så genetisk rådgivning anbefales, hvis du vil have børn.
Som med de andre trisomier udvikler trisomi X sig oftest som en fri trisomi, dvs. på grund af en mangel på opdeling (ikke-sammenhæng) af søsterchromatiderne. Også her opstår det normalt under modningen af de moderlige ægceller, skønt sandsynligheden øges med alderen.
Skørt X-syndrom
Fragilt X-syndrom eller Martin Bell-syndrom foretrækkes hos mænd, da de kun har et X-kromosom og derfor er mere påvirket af ændringen.
Det forekommer med en frekvens på 1: 1250 blandt de levende mandlige fødsler på et år, hvilket gør det til den mest almindelige form for uspecifik mental retardering, dvs. alle psykiske handicap, der ikke kan beskrives ved et specielt syndrom med typiske tegn.
Fragilt X-syndrom kan normalt også forekomme hos piger i en noget svagere form, hvilket skyldes utilsigtet inaktivering af et af X-kromosomerne. Jo højere andel af det slukkede sunde X-kromosom er, jo stærkere er symptomerne.
I de fleste tilfælde er kvinder imidlertid bærere af forudsigelsen, som endnu ikke giver nogen kliniske symptomer, men øger sandsynligheden for fuld mutation i deres sønner massivt. I meget sjældne tilfælde kan mænd også være bærere af forudsigelsen, som de derefter kun kan videregive til døtre, men som normalt også er klinisk sunde (Sherman-paradoks).
Syndromet udløses af et ekstremt forøget antal CGG-tripletter (en bestemt basesekvens) i FMR-genet (skrøbelige-sted-mental retardering); i stedet for 10-50 kopier, forudsætningen 50-200, med fuld udtryk 200-2000 eksemplarer.
Under lysmikroskopet ser det ud som et brud i den lange arm, hvilket er det, der gav syndromet sit navn. Dette fører til deaktivering af det berørte gen, hvilket igen forårsager symptomerne.
Berørte mennesker viser en langsommelig udvikling af tale og bevægelse og kan vise adfærdsmæssige problemer, der kan føre i retning af hyperaktivitet, men også autisme.Rent ydre abnormaliteter (tegn på dysmorfisme) er et langt ansigt med en fremtrædende hage og udstående ører. Med puberteten forstørres testiklerne meget kraftigt (makroorchidia), og ansigtstrækkene bliver grovere. Der er en let ophobning af psykologiske abnormiteter og en særlig tidlig overgangsalder blandt kvindelige bærere af premutationen.
Hvad er en kromosomanalyse?
Kromosomanalyse er en proces inden for cytogenetik, med hvilken numeriske eller strukturelle kromosomafvigelser kan påvises.
En sådan analyse ville f.eks. Blive anvendt, hvis et kromosomalt syndrom øjeblikkeligt er mistænkt, dvs. i tilfælde af misdannelser (dysmorfismer) eller intellektuel handicap (retardering), men også i tilfælde af infertilitet, regelmæssige aborter (aborter) og også med visse kræftformer (f.eks. Lymfomer) eller leukæmi).
Dette kræver normalt lymfocytter, en speciel type immuncelle, der opnås fra patientens blod. Da der kun kan opnås en relativt lille mængde på denne måde, stimuleres cellerne til at dele sig med phytohemagglutinin, og lymfocytterne kan derefter dyrkes i laboratoriet.
I nogle tilfælde udtages i stedet prøver (biopsier) fra huden eller rygmarven, og en lignende procedure anvendes. Målet er at få så meget DNA-materiale som muligt, der i øjeblikket er midt i celledelingen. I metafasen er alle kromosomer arrangeret i et niveau omtrent midt i cellen for at blive trukket mod de modsatte sider (poler) af cellen i det næste trin, anafasen.
På dette tidspunkt er kromosomerne særlig tæt pakket (stærkt kondenseret). Spindelfgiften colchicine tilsættes, som fungerer nøjagtigt i denne fase af cellecyklussen, så metafasekromosomerne akkumuleres. Derefter isoleres og farves de ved hjælp af specielle farvningsmetoder.
Det mest almindelige er GTG-banding, hvor kromosomerne behandles med trypsin, et fordøjelsesenzym og pigmentet Giemsa. De særligt tætpakkede regioner og regionerne rige på adenin og thymin er mørke.
De resulterende G-bånd er karakteristiske for hvert kromosom og anses forenklet for at være regionerne med færre gener. Et billede af kromosomerne farvet på denne måde er taget i tusindfoldig forstørrelse og der oprettes et karyogram ved hjælp af et computerprogram. Foruden båndmønsteret bruges kromosomets størrelse og centromerens placering til at hjælpe med at arrangere kromosomerne i overensstemmelse hermed. Der er også andre metoder til banding, der kan have meget forskellige fordele.
Anbefalinger fra redaktionen
Se følgende artikler for mere generel information:
- Cellekernedeling
- Funktioner af cellekernen
- Trisomi 21
- Genetiske sygdomme

.jpg)